- About
- Mission Statement
Education. Evidence. Regrowth.
- Education.
Prioritize knowledge. Make better choices.
- Evidence.
Sort good studies from the bad.
- Regrowth.
Get bigger hair gains.
Team MembersPhD's, resarchers, & consumer advocates.
- Rob English
Founder, researcher, & consumer advocate
- Research Team
Our team of PhD’s, researchers, & more
Editorial PolicyDiscover how we conduct our research.
ContactHave questions? Contact us.
Before-Afters- Transformation Photos
Our library of before-after photos.
- — Jenna, 31, U.S.A.
I have attached my before and afters of my progress since joining this group...
- — Tom, 30, U.K.
I’m convinced I’ve recovered to probably the hairline I had 3 years ago. Super stoked…
- — Rabih, 30’s, U.S.A.
My friends actually told me, “Your hairline improved. Your hair looks thicker...
- — RDB, 35, New York, U.S.A.
I also feel my hair has a different texture to it now…
- — Aayush, 20’s, Boston, MA
Firstly thank you for your work in this field. I am immensely grateful that...
- — Ben M., U.S.A
I just wanted to thank you for all your research, for introducing me to this method...
- — Raul, 50, Spain
To be honest I am having fun with all this and I still don’t know how much...
- — Lisa, 52, U.S.
I see a massive amount of regrowth that is all less than about 8 cm long...
Client Testimonials150+ member experiences.
 Scroll DownPopular Treatments
Scroll DownPopular Treatments- Treatments
Popular treatments. But do they work?
- Finasteride
- Oral
- Topical
- Dutasteride
- Oral
- Topical
- Mesotherapy
- Minoxidil
- Oral
- Topical
- Ketoconazole
- Shampoo
- Topical
- Low-Level Laser Therapy
- Therapy
- Microneedling
- Therapy
- Platelet-Rich Plasma Therapy (PRP)
- Therapy
- Scalp Massages
- Therapy
- More
IngredientsTop-selling ingredients, quantified.
- Saw Palmetto
- Redensyl
- Melatonin
- Caffeine
- Biotin
- Rosemary Oil
- Lilac Stem Cells
- Hydrolyzed Wheat Protein
- Sodium Lauryl Sulfate
- More
ProductsThe truth about hair loss "best sellers".
- Minoxidil Tablets
Xyon Health
- Finasteride
Strut Health
- Hair Growth Supplements
Happy Head
- REVITA Tablets for Hair Growth Support
DS Laboratories
- FoliGROWTH Ultimate Hair Neutraceutical
Advanced Trichology
- Enhance Hair Density Serum
Fully Vital
- Topical Finasteride and Minoxidil
Xyon Health
- HairOmega Foaming Hair Growth Serum
DrFormulas
- Bio-Cleansing Shampoo
Revivogen MD
- more
Key MetricsStandardized rubrics to evaluate all treatments.
- Evidence Quality
Is this treatment well studied?
- Regrowth Potential
How much regrowth can you expect?
- Long-Term Viability
Is this treatment safe & sustainable?
Free Research- Free Resources
Apps, tools, guides, freebies, & more.
- Free CalculatorTopical Finasteride Calculator
- Free Interactive GuideInteractive Guide: What Causes Hair Loss?
- Free ResourceFree Guide: Standardized Scalp Massages
- Free Course7-Day Hair Loss Email Course
- Free DatabaseIngredients Database
- Free Interactive GuideInteractive Guide: Hair Loss Disorders
- Free DatabaseTreatment Guides
- Free Lab TestsProduct Lab Tests: Purity & Potency
- Free Video & Write-upEvidence Quality Masterclass
- Free Interactive GuideDermatology Appointment Guide
- More
Articles100+ free articles.
-
Which Hormones Cause Hair Loss in Females?
-
Does Zepbound Cause Hair Loss?
-
Does Wegovy Cause Hair Loss?
-
Strut Health Review: 5 Things to Consider Before Purchasing
-
Does Tirzepatide Cause Hair Loss?
-
Musely Reviews for Hair Loss: Are They Trustworthy?
-
Best Minoxidil for Women: Top 6 Brands of 2026
-
Best Topical Finasteride: 5 Top Products of 2026
PublicationsOur team’s peer-reviewed studies.
- Microneedling and Its Use in Hair Loss Disorders: A Systematic Review
- Use of Botulinum Toxin for Androgenic Alopecia: A Systematic Review
- Conflicting Reports Regarding the Histopathological Features of Androgenic Alopecia
- Self-Assessments of Standardized Scalp Massages for Androgenic Alopecia: Survey Results
- A Hypothetical Pathogenesis Model For Androgenic Alopecia:Clarifying The Dihydrotestosterone Paradox And Rate-Limiting Recovery Factors
Menu- AboutAbout
- Mission Statement
Education. Evidence. Regrowth.
- Team Members
PhD's, resarchers, & consumer advocates.
- Editorial Policy
Discover how we conduct our research.
- Contact
Have questions? Contact us.
- Before-Afters
Before-Afters- Transformation Photos
Our library of before-after photos.
- Client Testimonials
Read the experiences of members
Before-Afters/ Client Testimonials- Popular Treatments
-
ArticlesWhich Hormones Cause Hair Loss in Females?
First Published Jun 30 2026Last Updated Jun 30 2026Miscellaneous Researched & Written By:Sarah King, PhD
Researched & Written By:Sarah King, PhD Reviewed By:Rob English, Medical Editor
Reviewed By:Rob English, Medical Editor
Want help with your hair regrowth journey?
Get personalized support, product recommendations, video calls, and more from our researchers, trichologists, and PhD's dedicated to getting you the best possible outcomes.
Learn MoreArticle Summary
Hair loss in women isn’t driven by a single hormone or pathway. While androgens like DHT play a role in some cases, factors such as estrogen decline, adrenal hormones, prolactin, and chronic stress can all disrupt normal hair cycling. This article explores the evidence behind each hormone and explains why female hair thinning is often multifactorial rather than purely androgen‑driven.
Full Article
Female pattern hair loss (FPHL) is the most common cause of hair thinning in women. Unlike male androgenic alopecia (AGA), where dihydrotestosterone (DHT) is the main driver, FPHL appears to involve a more complex interplay of hormones, not all of them androgens. So, what do we know about the hormones involved in FPHL, and which ones are they? We will walk you through the current evidence for each implicated hormone and explain what the evidence says about the role of each one in hair loss.
Is Female Pattern Hair Loss Just “Female AGA”?
Before we discuss individual hormones, it is worth asking a more fundamental question: Is FPHL actually the same condition as male AGA, just presenting in a different body?
For a long time, the assumption was yes. Both conditions involve progressive follicle miniaturization, where individual hairs become progressively finer, shorter, and less pigmented over time until the follicle eventually stops producing a visible hair altogether. Both are influenced by genetics, and both have historically been grouped under the term AGA, implying androgens as the causal mechanism.
But a growing body of evidence suggests that this grouping may have been premature. In men, the case for DHT as the primary driver of AGA is strong across three main areas of evidence:
- Mechanistic: When AGA-prone hair follicles are exposed to DHT in the lab, dermal papilla cells undergo apoptosis (programmed cell death).[1]Randall, V.A., Hibberts, N.A., Hamada, K. (1996). A comparison of the culture and growth of dermal papilla cells from hair follicles from non-balding and balding (androgenetic alopecia) scalp. … Continue reading
- Observational: Men who lack the gene to produce type II 5ɑ-reductase (the enzyme that converts testosterone to DHT) do not develop pattern baldness in adulthood. Furthermore, men castrated before puberty, who reduce their exposure to male hormones by approximately 95%, also do not go bald.[2]Imperato-McGinley, J., Guerrero, L., Gautier, T., Peterson, R.E. (1974). Steroid 5alpha-reductase deficiency in man: an inherited form of male pseudohermaphroditism. Science. 186(4170). 1213–1215. … Continue reading
- Interventional: DHT‑reducing drugs such as finasteride and dutasteride stabilize or improve hair in the great majority of men with AGA, with randomized and long‑term studies showing no further visible hair loss in approximately 80% or more of treated men over 2–5 years.[3]McClellan, K.J., Markham, A. (1999). Finasteride: a review of its use in male pattern hair loss. Drugs. 57(1). 111-126. Available at: https://doi.org/10.2165/00003495-199957010-00014,[4]Vano-Galvan, S., Saceda-Corralo, D., Moreno-Arrones, O.M., Rodrigues-Barata, R., Morales, C., Gil-Redondo, R., Bernardez-Guerra, C., Hermosa-Gelbard, A., Jaen-Olasolo, P. (2019). Effectiveness and … Continue reading
This creates a compelling case for DHT as the primary driver in male AGA. But when we apply the same framework to women, the picture becomes far more complicated.
Let’s think about the following:
- Women with complete androgen insensitivity syndrome (CAIS), a condition in which a person carries a mutation that renders androgen receptors completely non-functional, can still develop FPHL, despite having no androgenic activity anywhere in the body.[5]Cousen, P., Messenger, A. (2010). Female pattern hair loss in complete androgen insensitivity syndrome. British Journal of Dermatology. 162(5). 1135–1137. Available at: … Continue reading
- Most women with FPHL have completely normal androgen levels on blood testing.[6]Orme, S., Cullen, D.R., Messenger, A.G. (1999). Diffuse female hair loss: are androgens necessary? British Journal of Dermatology. 141(3). 521–523. Available at: … Continue reading
- Finasteride can fail to improve hair outcomes in a proportion of women with FPHL. For example, a randomized controlled trial of 67 postmenopausal women with normal testosterone showed that 1 mg finasteride reduced serum DHT but had no measurable effect on hair loss compared to a placebo.[7]Price, V.H., Roberts, J.L., Hordinsky, M., Olsen, E.A., Savin, R., Bergfeld, W., Fiedler, V., Lucky, A., Whiting, D.A., Pappas, F., Culbertson, J., Kotey, P., Meehan, A., Waldstreicher, J. (2000). … Continue reading
- In androgen-deficient women, testosterone replacement therapy has been associated with improved hair outcomes. In a questionnaire study of 285 women treated for androgen deficiency, 76 (27%) reported hair thinning before treatment, of whom 63% reported hair growth on testosterone therapy. Importantly, no patient reported hair loss as a result of the treatment.[8]Glaser, R.L., Dimitrakakis, C., Messenger, A.G. (2012). Improvement in scalp hair growth in androgen-deficient women treated with testosterone: a questionnaire study. British Journal of Dermatology. … Continue reading
These are not minor anomalies. They suggest that factors outside of DHT and possibly outside of androgens altogether play a meaningful role in many cases of FPHL. Clinicians have begun to acknowledge that male AGA and FPHL may constitute distinct subtypes of genetically mediated hair loss that were perhaps never best grouped together.[9]Brough, K.R., Torgerson, R.R. (2017). Hormonal therapy in female pattern hair loss. International Journal of Women’s Dermatology. 3(1). 53–57. Available at: … Continue reading
With that said, let’s look at what the evidence says for DHT and testosterone.
DHT and Testosterone – Still the Primary Suspects
Despite the above, androgens remain the best-evidenced candidates for a significant proportion of FPHL cases. The question is not whether androgens are involved, but which women, and which androgens.
DHT
DHT is produced when the enzyme 5ɑ-reductase converts free testosterone into a more potent androgenic form. Both type I and type II isoforms of 5ɑ-reductase are expressed in scalp hair follicles, and DHT’s binding to androgen receptors in dermal papilla cells triggers the miniaturization process, shortening the anagen (growth) phase and progressively shrinking follicle diameter.
The clearest evidence that DHT drives FPHL in women comes from two sources.
First, women with conditions involving androgen excess, most notably polycystic ovary syndrome (now known as Polyendocrine Metabolic Ovarian Syndrome – PMOS), show elevated 5ɑ-reductase activity, producing more DHT peripherally and experiencing FPHL at higher rates. A 2017 systematic review and meta-analysis confirmed significantly elevated 5ɑ-reductase in women with PMOS compared to controls.[10]Wu, C., Wei, K., Jiang, Z. (2017). 5α-reductase activity in women with polycystic ovary syndrome: a systematic review and meta-analysis. Reproductive Biology and Endocrinology. 15. Article 21. … Continue reading
Second, androgen receptor-blocking drugs produce meaningful hair improvements in women with FPHL. For example, oral antiandrogens such as spironolactone or cyproterone acetate halt progression or induce visible regrowth in approximately 88% of treated women, and flutamide has been associated with improvement in over 80% of cases in prospective cohorts.[11]Sinclair, R., Wewerinke, M., Jolley D. (2005). Treatment of female pattern hair loss with oral antiandrogens. British Journal of Dermatology. 152(3). 466-473. Available at: … Continue reading,[12]Paradisi, R., Porcu, E., Fabbri, R., Seracchioli, R., Battaglia, C., Venturoli, S. (2011). Prospective cohort study on the effects and tolerability of flutamide in patients with female pattern hair … Continue reading
The FPHL population where DHT is most clearly implicated is relatively specific: women with co-existing androgen excess, those with PMOS, congenital adrenal hyperplasia (CAH), elevated free androgen index, or those whose hair loss demonstrably responds to finasteride therapy. In the majority of women with FPHL who have normal androgen levels, the role of DHT is less clear.
Testosterone
Testosterone itself also binds directly to androgen receptors in dermal papilla cells. Its relationship with female hair is, however, genuinely paradoxical.
On one hand, excess testosterone (as in PMOS or exogenous steroid use) is associated with FPHL. On the other hand, the testosterone questionnaires study we mentioned above found that 63% of androgen-deficient women with hair thinning reported hair growth after testosterone therapy, and none of the 285 women reported hair loss during treatment.
This suggests that in low androgen states, restoring testosterone may be net positive for hair health, or that the relative balance between androgens and estrogens matters more than absolute testosterone values.
One important thing should be noted about studies using blood tests for testosterone and DHT is that they may not capture the full picture.
A 2026 cross-sectional study measured serum levels of 3ɑ-androstanediol glucuronide (3ɑ-diol G), a metabolite that reflects how much DHT is being produced and metabolized within tissues, in 44 women with FPHL and 30 controls. Women with FPHL had significantly higher 3ɑ-diol G levels, and levels correlated positively with FPHL severity even when standard androgen markers like total testosterone and DHEA-S were within the normal range. Furthermore, analysis showed a high diagnostic accuracy for 3ɑ-diol G (82% sensitivity, 73% specificity).[13]Kathirvelu, A.P., Sardana, K., Sharma, L., Khurana, A., Sharath, S. (2026). Evidence of Increased 3-Alpha-Androstanediol Glucuronide in Female Pattern Alopecia: A Cross-Sectional Observational Study … Continue reading
This is a meaningful finding, suggesting that peripheral tissue DHT metabolism may be elevated in women with FPHL even in the absence of detectable systemic hyperandrogenism, which would help explain why many people with “ normal” blood results still appear to benefit from anti-androgen therapy.
DHEA-S and the Adrenal Androgen Connection
DHEA-S (dehydroepiandrosterone sulfate) is produced almost exclusively by the adrenal glands, not the ovaries, and serves as a circulating precursor that can be converted into testosterone and DHT in peripheral tissues.[14]Lin, H-Y., Chen, J-H., Chen, K-H. (2025). The Sex Hormone Precursors Dehydroepiandrosterone (DHEA) and Its Sulfate Ester Form (DHEAS): Molecular Mechanisms and Actions on Human Body. International … Continue reading
Its role in FPHL becomes particularly important when you consider what happens with flutamide, a non-selective androgen receptor blocker. Unlike finasteride, which specifically targets DHT production, flutamide blocks androgen receptor activity for all androgens, including testosterone, DHEA and its derivatives, and has been shown to specifically reduce adrenal androgen output.
One study investigated flutamide’s effect on pituitary and adrenal responsiveness to corticotropin-releasing factor (CRF) and found that it caused a significant reduction in adrenal androgen response, demonstrating that its mechanism of action extends beyond the ovaries to the adrenal glands.[15]De Leo, V., la Marca, A., Lanzetta, D., Cariello, P.L., D’Antona, D., Morgante, G. (1998). Effects of flutamide on pituitary and adrenal responsiveness to corticotropin-releasing factor (CRF). … Continue reading
In a randomized controlled trial of 48 hyperandrogenic premenopausal women with FPHL, patients were assigned to one of four groups: flutamide 250 mg/day, finasteride 5 mg/day, cyproterone acetate with ethinyl estradiol, or no treatment, for one year.[16]Carmina, E., Lobo, R.A. (2003). Treatment of hyperandrogenic alopecia in women. Fertility and Sterility. 79(1). 91–95. Available at: https://doi.org/10.1016/S0015-0282(02)04551-X Flutamide was the only agent that produced a statistically significant reduction in Ludwig scores, a 21% decrease. Neither finasteride nor cyproterone acetate produced significant changes in Ludwig score compared to the untreated control group. Patient and investigator assessments followed the same trend: flutamide provided the greatest clinical response in terms of slowing hair loss.
The fact that broad androgen receptor blockade outperformed DHT-specific inhibition in FPHL points toward a role for non-DHT androgens, of which DHEA-S is the primary adrenal precursor, in driving follicle miniaturization in at least some women.
The adrenal connection also explains why FPHL continues and often worsens after menopause, when ovarian androgen production has effectively ceased. The adrenal glands become the dominant androgen source, and even though absolute DHEA-S levels decline with age (a process called adrenopause), longitudinal data showed that several adrenal androgens, including DHEA and androstenediol, actually rise during the menopausal transition in most women, before eventually declining in late menopause.[17]Lasley, B.L., McConnell, D.S., Crawford, S. (2011). Adrenal androgens and the menopausal transition. Obstetrics and Gynecology Clinics of North America. 38(3). 467–475. Available at: … Continue reading The result is that even without any absolute rise in androgen values, the androgen-to-estrogen ratio at the hair follicle increases significantly, creating a follicular environment that is relatively more androgenic precisely at the life stage when FPHL tends to emerge or accelerate.[18]Trink, A., Pinto, D., Giuliani, G., Mondadori, G., Rinaldi, F. (2023). The Menopausal Transition: Is the Hair Follicle “Going through Menopause”? Biomedicines. 11(11). 3041. Available at: … Continue reading
Estradiol
Estradiol (E2), the primary and most potent estrogen in reproductive-age women, acts through estrogen receptor beta (ER-ꞵ), which is expressed in the hair follicle epithelium. Its role in hair biology is primarily protective and anagen-promoting.
Several sources of evidence support estradiol’s hair-preserving effects.
Pregnancy
Elevated estrogen during pregnancy is associated with a prolonged anagen phase and thicker, fuller-feeling hair. The sharp postpartum drop in estrogen then synchronizes a large number of follicles into telogen simultaneously, producing the well-recognized postpartum telogen effluvium.[19]Piérard-Franchimont, C., Piérard, G.E. (2013). Alterations in hair follicle dynamics in women. BioMed Research International. 2013. 957432. Available at: https://doi.org/10.1155/2013/957432 This is a real-world demonstration of estrogen’s direct influence on hair cycle timing.
Menopause
A cross-sectional study of 178 postmenopausal women found an FPHL prevalence of 52.2%, with age and time since menopause both significantly associated.[20]Chaikittisilpa, S., Rattanasirisin, N., Panchaprateep, R., Orprayoon, N., Phutrakul, P., Suwan, A., Jaisamrarn, U. (2022). Prevalence of female pattern hair loss in postmenopausal women: a … Continue reading This indicates that FPHL rates rise markedly after menopause, precisely when ovarian estrogen production ceases.
Luteinizing Hormone (LH)
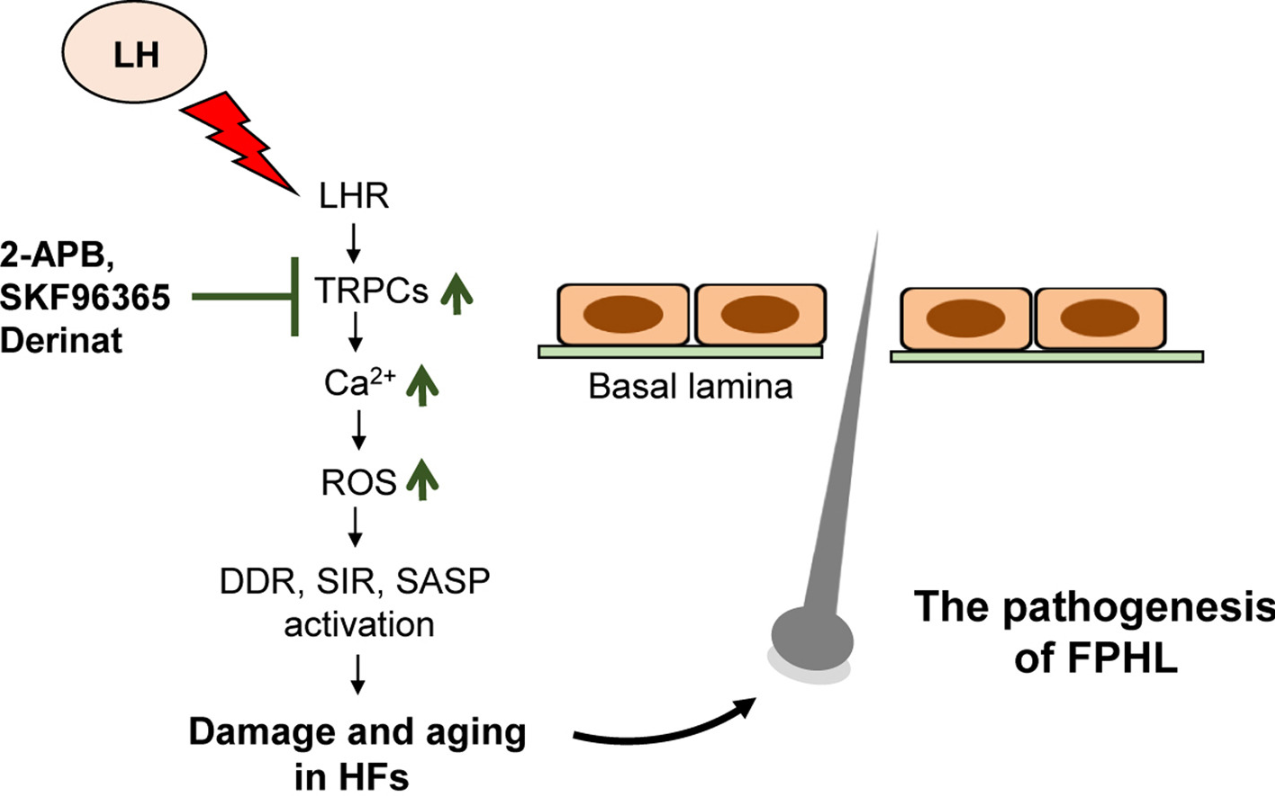
Emerging data on LH add another androgen-independent mechanism. As estrogen declines during menopause, LH levels surge. A 2025 study conducted in mice found a significant association between elevated LH and FPHL, with LH binding to receptors on dermal papilla cells activating downstream transient potential canonical (TRPC) channels, leading to increasing reactive oxygen species, elevated calcium signaling, senescence, and DNA damage response activation.[21]Wu, C.Y., Vadhan, A., Wei, W.Y., Yu, H.S., Hsieh, Y.C., Chen, W.C., Hsu, C.C., Liu, Y.H., Hsu, W.L. (2025). Luteinizing Hormone Induces Murine Hair Loss through Transient Receptor Potential Canonical … Continue reading This effectively accelerates hair follicle aging through a pathway entirely separate from androgens.

Figure 1: How luteinizing hormone (LH) may contribute to female pattern hair loss. After menopause, levels of LH rise significantly as estrogen declines. This figure shows how elevated LH may damage hair follicles through a chain reaction: LH binds to receptors on the cells that control hair growth (dermal papilla cells), triggering a cascade that floods those cells with calcium and generates harmful molecules called reactive oxygen species (ROS). This chemical stress activates the cell’s own damage-response systems, accelerating aging and dysfunction within the follicle. The result is a hair follicle that is less able to sustain normal growth, contributing to the pattern of thinning seen in FPHL. Importantly, this pathway operates entirely independently of androgens like DHT, which may help explain why some women develop FPHL without any detectable hormonal abnormality on standard blood tests.[22]Wu, C.Y., Vadhan, A., Wei, W.Y., Yu, H.S., Hsieh, Y.C., Chen, W.C., Hsu, C.C., Liu, Y.H., Hsu, W.L. (2025). Luteinizing Hormone Induces Murine Hair Loss through Transient Receptor Potential Canonical … Continue reading Image used in conjunction with the Creative Commons CC-BY License.
Overall, the decline in estradiol at menopause appears to contribute to FPHL both directly through the loss of anagen-promoting signaling and indirectly through the resulting shift in the androgen-to-estrogen ratio and the surge in LH that drives senescence via a separate oxidative stress pathway.
Prolactin
Prolactin (PRL) is best known as the hormone that triggers lactation. But its role in hair biology is more interesting than most people realize.
Human scalp hair follicles are both a target and source of PRL. The follicle itself produces prolactin locally and responds to it in an autocrine and/or paracrine manner.[23]Foitzik, K., Krause, K., Conrad, F., Nakamura, M., Funk, W., Paus, R. (2006). Human scalp hair follicles are both a target and a source of prolactin, which serves as an autocrine and/or paracrine … Continue reading This means that PRL is not only reaching the hair follicle through the bloodstream, but is also being produced and acted upon within the follicle itself.
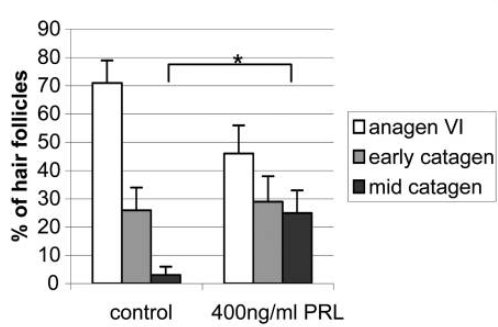
So what does prolactin actually do to the hair follicle? A study exposed organ-cultured human scalp hair follicles to 400 ng/ml of prolactin for 8 days.[24]Foitzik, K., Krause, K., Conrad, F., Nakamura, M., Funk, W., Paus, R. (2006). Human scalp hair follicles are both a target and a source of prolactin, which serves as an autocrine and/or paracrine … Continue reading This is the concentration seen in patients with hyperprolactinemia-associated TE. The results were as follows:
- Hair shaft elongation was significantly reduced: control follicles grew approximately 1.6 mm over 8 days, while prolactin-treated hair follicles grew only 1.3 mm.
- In control follicles, more than 70% remained in anagen after 8 days. In the prolactin-treated group, only 45% remained in anagen, while 25% had entered mid-catagen (regression phase), compared to only 3% in controls.
- Ki-67 (a marker of cell proliferation) was significantly lower in prolactin-treated follicles (47% vs 59% of proliferating cells; p<0.05).
- TUNEL-positive apoptotic cells were significantly increased in prolactin-treated follicles (p<0.01).

Figure 2: Effect of 400 ng/mL PRL on hair follicle cycle stage in isolated human hair follicles.[25]Foitzik, K., Krause, K., Conrad, F., Nakamura, M., Funk, W., Paus, R. (2006). Human scalp hair follicles are both a target and a source of prolactin, which serves as an autocrine and/or paracrine … Continue reading Image used in conjunction with the PMC Copyright Notice.
Importantly, the study also found that prolactin and its receptor are more intensely expressed in catagen follicles than in anagen follicles, suggesting prolactin may function as a natural catagen-inducing signal that becomes dysregulated at pathologically high levels.
The clinical picture is more nuanced, however. Normal physiological prolactin levels in non-pregnant women are approximately 5-20 ng/mL. The concentrations used in the above study (200-400 ng/mL) are only typically seen in women with prolactinomas (pituitary tumors producing excess prolactin) or those taking dopamine-antagonist medications. At these extreme levels, hair loss is a recognized clinical consequence. At modest elevations, the evidence is less clear, however.
Prolactin measurement is nonetheless a standard component of the hormonal workup for women presenting with diffuse hair loss.
Cortisol and the Stress-Hair Loss Axis
The hormones discussed so far are primarily relevant to FPHL. But now we will focus on TE, in which a triggering event causes a large number of follicles to simultaneously shift into the telogen (resting) phase, producing a wave of shedding 2-4 months later. TE has its own hormonal driver: cortisol.
Cortisol is the primary output of the hypothalamic-pituitary-adrenal (HPA) axis under stress. When the hypothalamus detects a stressor, it releases corticotropin-releasing hormone (CRH), which triggers the pituitary to release adrenocorticotropic hormone (ACTH), which in turn drives the adrenal glands to produce cortisol.[26]Dunlavey, C.J. (2018). Introduction to the Hypothalamic-Pituitary-Adrenal Axis: Healthy and Dysregulated Stress Responses, Developmental Stress and Neurodegeneration. Journal of Undergraduate … Continue reading
Here is what makes this particularly relevant to hair biology: the hair follicle has its own functional, local HPA axis.[27]Ito, N., Ito, T., Kromminga, A., Bettermann, A., Takigawa, M., Kees, F., Straub, R.H., Paus, R. (2005). Human hair follicles display a functional equivalent of the hypothalamic-pituitary-adrenal axis … Continue reading
Dermal papilla cells express CRH, ACTH, cortisol, and their corresponding receptors, meaning the follicle can mount a stress response locally, independently of the systemic circulation. When this local axis is activated, CRH inhibits hair shaft elongation, triggers premature catagen transition, and increases oxidative stress and reactive oxygen species.
The most mechanistically compelling pathway identified to date involves a protein called Gas6. Gas6 is a signaling molecule produced by dermal papilla cells that activates hair follicle stem cells out of their quiescent (resting) state and into a new anagen cycle.
A landmark 2021 study in Nature demonstrated that chronic elevation of corticosterone (the rodent equivalent of human cortisol) suppresses Gas6 expression in dermal papilla cells, which effectively locks stem cells in extended telogen and prevents transition to a new anagen phase. [28]Choi, S., Zhang, B., Ma, S., Gonzalez-Celeiro, M., Stein, D., Jin, X., Kim, S.T., Kang, Y.L., Besnard, A., Rezza, A., Grisanti, L., Buenrostro, J.D., Rendl, M., Nahrendorf, M., Sahay, A., Hsu, Y.C. … Continue reading In mice with no adrenal glands (and therefore no cortisol), hair cycling was normalized without intervention. Restoring corticosterone via oral supplementation reproduced the telogen-prolonging effect.
Restoring Gas6 expression via adenovirus-associated injection was sufficient to reactivate HFSCs and promote hair growth even in a high-corticosterone environment.
The implication is clear: chronically elevated cortisol can directly suppress the molecular signal that reactivates follicle stem cells and initiates a new growth cycle.
At a Glance: Hormones Implicated in Female Hair Loss
Hormone Condition Mechanism Strength of Evidence DHT FPHL (androgen-excess subtype) AR-mediated follicle miniaturization via dermal papilla apoptosis Strong in hyperandrogenic women (PCOS, CAH) Testosterone FPHL Direct AR binding; peripheral conversion to DHT via 5ɑ-reductase Moderate – paradoxical in normo/hypoandrogenic women DHEA-S FPHL (including postmenopausal) Adrenal androgen precursor; peripheral DHT synthesis independent of ovarian input Moderate – supported by flutamide’s superiority over finasteride Estradiol FPHL Anagen-prolonging; aromatase activity protects the frontal hairline; declines sharply at menopause relative to androgens Moderate – epidemiological and mechanistic data LH FPHL (postmenopausal) TRPC-mediated oxidative stress and cellular senescence in dermal papilla cells Emerging – 2024 data only PRL TE; FPHL (hyperprolactinaemia) Local catagen-promoting signal in the follicle at pathologically elevated levels Emerging – in mice only, clear at high levels, modest elevations inconclusive Cortisol TE Gas6 suppression leads to HFSC quiescence and extended telogen phase Moderate – strong mechanistic data in rodents; human data building Final Thoughts
Female hair loss is rarely the result of a single hormone acting in isolation. For many women, the androgen story (DHT, testosterone, and adrenal-derived DHEA-S) remains central, but it is incomplete on its own. Estradiol withdrawal at menopause, surging LH, elevated prolactin, and chronic cortisol each represent distinct biological pathways capable of driving or accelerating hair loss through mechanisms that have nothing to do with androgens. What this means practically is that two women presenting with identical hairline photographs may need different interventions, and that a thorough hormonal workup is probably the most logical starting point.
References[+]
References ↑1 Randall, V.A., Hibberts, N.A., Hamada, K. (1996). A comparison of the culture and growth of dermal papilla cells from hair follicles from non-balding and balding (androgenetic alopecia) scalp. British Journal of Dermatology. 134(3). 437–444. Available at: https://doi.org/10.1111/j.1365-2133.1996.tb16224.x ↑2 Imperato-McGinley, J., Guerrero, L., Gautier, T., Peterson, R.E. (1974). Steroid 5alpha-reductase deficiency in man: an inherited form of male pseudohermaphroditism. Science. 186(4170). 1213–1215. Available at: https://doi.org/10.1126/science.186.4170.1213 ↑3 McClellan, K.J., Markham, A. (1999). Finasteride: a review of its use in male pattern hair loss. Drugs. 57(1). 111-126. Available at: https://doi.org/10.2165/00003495-199957010-00014 ↑4 Vano-Galvan, S., Saceda-Corralo, D., Moreno-Arrones, O.M., Rodrigues-Barata, R., Morales, C., Gil-Redondo, R., Bernardez-Guerra, C., Hermosa-Gelbard, A., Jaen-Olasolo, P. (2019). Effectiveness and safety of oral dutasteride for male androgenetic alopecia in real clinical practice: a descriptive monocentric study. Dermatologic Therapy. 33(1). E13182. Available at: https://doi.org/10.1111/dth.13182 ↑5 Cousen, P., Messenger, A. (2010). Female pattern hair loss in complete androgen insensitivity syndrome. British Journal of Dermatology. 162(5). 1135–1137. Available at: https://doi.org/10.1111/j.1365-2133.2010.09661.x ↑6 Orme, S., Cullen, D.R., Messenger, A.G. (1999). Diffuse female hair loss: are androgens necessary? British Journal of Dermatology. 141(3). 521–523. Available at: https://doi.org/10.1046/j.1365-2133.1999.03049.x ↑7 Price, V.H., Roberts, J.L., Hordinsky, M., Olsen, E.A., Savin, R., Bergfeld, W., Fiedler, V., Lucky, A., Whiting, D.A., Pappas, F., Culbertson, J., Kotey, P., Meehan, A., Waldstreicher, J. (2000). Lack of efficacy of finasteride in postmenopausal women with androgenetic alopecia. Journal of the American Academy of Dermatology. 43(5). 768-776. Available at: https://doi.org/10.1067/mjd.2000.107953 ↑8 Glaser, R.L., Dimitrakakis, C., Messenger, A.G. (2012). Improvement in scalp hair growth in androgen-deficient women treated with testosterone: a questionnaire study. British Journal of Dermatology. 166(2). 274–278. Available at: https://doi.org/10.1111/j.1365-2133.2011.10655.x ↑9 Brough, K.R., Torgerson, R.R. (2017). Hormonal therapy in female pattern hair loss. International Journal of Women’s Dermatology. 3(1). 53–57. Available at: https://doi.org/10.1016/j.ijwd.2017.01.001 ↑10 Wu, C., Wei, K., Jiang, Z. (2017). 5α-reductase activity in women with polycystic ovary syndrome: a systematic review and meta-analysis. Reproductive Biology and Endocrinology. 15. Article 21. Available at: https://doi.org/10.1186/s12958-017-0242-9 ↑11 Sinclair, R., Wewerinke, M., Jolley D. (2005). Treatment of female pattern hair loss with oral antiandrogens. British Journal of Dermatology. 152(3). 466-473. Available at: https://doi.org/10.1111/j.1365-2133.2005.06218.x. ↑12 Paradisi, R., Porcu, E., Fabbri, R., Seracchioli, R., Battaglia, C., Venturoli, S. (2011). Prospective cohort study on the effects and tolerability of flutamide in patients with female pattern hair loss. The Annals of Pharmacotherapy. 45(4). 469-475. Available at: https://doi.org/10.1345/aph.1P600 ↑13 Kathirvelu, A.P., Sardana, K., Sharma, L., Khurana, A., Sharath, S. (2026). Evidence of Increased 3-Alpha-Androstanediol Glucuronide in Female Pattern Alopecia: A Cross-Sectional Observational Study Evaluating Ovarian, Adrenal, and Peripheral Androgens. International Journal of Dermatology. Available at: https://doi.org/10.1111/ijd.70445 ↑14 Lin, H-Y., Chen, J-H., Chen, K-H. (2025). The Sex Hormone Precursors Dehydroepiandrosterone (DHEA) and Its Sulfate Ester Form (DHEAS): Molecular Mechanisms and Actions on Human Body. International Journal of Molecular Sciences. 26(17). 8568. Available at: https://doi.org/10.3390/ijms26178568 ↑15 De Leo, V., la Marca, A., Lanzetta, D., Cariello, P.L., D’Antona, D., Morgante, G. (1998). Effects of flutamide on pituitary and adrenal responsiveness to corticotropin-releasing factor (CRF). Clinical Endocrinology. 49(1). 85–89. Available at: https://doi.org/10.1046/j.1365-2265.1998.00483.x ↑16 Carmina, E., Lobo, R.A. (2003). Treatment of hyperandrogenic alopecia in women. Fertility and Sterility. 79(1). 91–95. Available at: https://doi.org/10.1016/S0015-0282(02)04551-X ↑17 Lasley, B.L., McConnell, D.S., Crawford, S. (2011). Adrenal androgens and the menopausal transition. Obstetrics and Gynecology Clinics of North America. 38(3). 467–475. Available at: https://doi.org/10.1016/j.ogc.2011.06.001 ↑18 Trink, A., Pinto, D., Giuliani, G., Mondadori, G., Rinaldi, F. (2023). The Menopausal Transition: Is the Hair Follicle “Going through Menopause”? Biomedicines. 11(11). 3041. Available at: https://doi.org/10.3390/biomedicines11113041 ↑19 Piérard-Franchimont, C., Piérard, G.E. (2013). Alterations in hair follicle dynamics in women. BioMed Research International. 2013. 957432. Available at: https://doi.org/10.1155/2013/957432 ↑20 Chaikittisilpa, S., Rattanasirisin, N., Panchaprateep, R., Orprayoon, N., Phutrakul, P., Suwan, A., Jaisamrarn, U. (2022). Prevalence of female pattern hair loss in postmenopausal women: a cross-sectional study. Menopause. 29(4). 415–420. Available at: https://doi.org/10.1097/GME.0000000000001927 ↑21, ↑22 Wu, C.Y., Vadhan, A., Wei, W.Y., Yu, H.S., Hsieh, Y.C., Chen, W.C., Hsu, C.C., Liu, Y.H., Hsu, W.L. (2025). Luteinizing Hormone Induces Murine Hair Loss through Transient Receptor Potential Canonical Channel-Mediated Cell Aging Responses: Implications for Female Pattern Hair Loss Pathogenesis. Journal of Investigative Dermatology. Available at: https://doi.org/10.1016/j.jid.2025.03.044 ↑23, ↑25 Foitzik, K., Krause, K., Conrad, F., Nakamura, M., Funk, W., Paus, R. (2006). Human scalp hair follicles are both a target and a source of prolactin, which serves as an autocrine and/or paracrine promoter of apoptosis-driven hair follicle regression. American Journal of Pathology. 168(3). 748–756. Available at: https://doi.org/10.2353/ajpath.2006.050468 ↑24 Foitzik, K., Krause, K., Conrad, F., Nakamura, M., Funk, W., Paus, R. (2006). Human scalp hair follicles are both a target and a source of prolactin, which serves as an autocrine and/or paracrine promoter of apoptosis-driven hair follicle regression. American Journal of Pathology. 168(3). 748–756. Available at: https://doi.org/10.2353/ajpath.2006.050468 ↑26 Dunlavey, C.J. (2018). Introduction to the Hypothalamic-Pituitary-Adrenal Axis: Healthy and Dysregulated Stress Responses, Developmental Stress and Neurodegeneration. Journal of Undergraduate Neuroscience Education. 16(2). R59-R60. Available at: PMID: 30057514 ↑27 Ito, N., Ito, T., Kromminga, A., Bettermann, A., Takigawa, M., Kees, F., Straub, R.H., Paus, R. (2005). Human hair follicles display a functional equivalent of the hypothalamic-pituitary-adrenal axis and synthesize cortisol. FASEB J. 19(10). 1332-1334. Available at: https://doi.org/10.1096/fj.04-1968fje. ↑28 Choi, S., Zhang, B., Ma, S., Gonzalez-Celeiro, M., Stein, D., Jin, X., Kim, S.T., Kang, Y.L., Besnard, A., Rezza, A., Grisanti, L., Buenrostro, J.D., Rendl, M., Nahrendorf, M., Sahay, A., Hsu, Y.C. (2021). Corticosterone inhibits GAS6 to govern hair follicle stem-cell quiescence. Nature. 592(7854). 428–432. Available at: https://doi.org/10.1038/s41586-021-03417-2 Want help with your hair regrowth journey?
Get personalized support, product recommendations, video calls, and more from our researchers, trichologists, and PhD's dedicated to getting you the best possible outcomes.
Learn More
Sarah King, PhD
Dr. Sarah King is a researcher & writer who holds a BSc in Medical Biology, an MSc in Forensic Biology, and a Ph.D. in Molecular and Cellular Biology. While at university, Dr. King’s research focused on cellular aging and senescence through NAD-dependent signaling – along with research into prostaglandins and their role in hair loss. She is a co-author on several upcoming manuscripts with the Perfect Hair Health team.
"... Can’t thank @Rob (PHH) and @sanderson17 enough for allowing me to understand a bit what was going on with me and why all these [things were] happening ... "
— RDB, 35, New York, U.S.A."... There is a lot improvement that I am seeing and my scalp feel alive nowadays... Thanks everyone. "
— Aayush, 20’s, Boston, MA"... I can say that my hair volume/thickness is about 30% more than it was when I first started."
— Douglas, 50’s, Montréal, CanadaWant help with your hair regrowth journey?
Get personalized support, product recommendations, video calls, and more from our researchers, trichologists, and PhD's dedicated to getting you the best possible outcomes.
Join Now - Mission Statement
 Scroll Down
Scroll Down More
More More
More